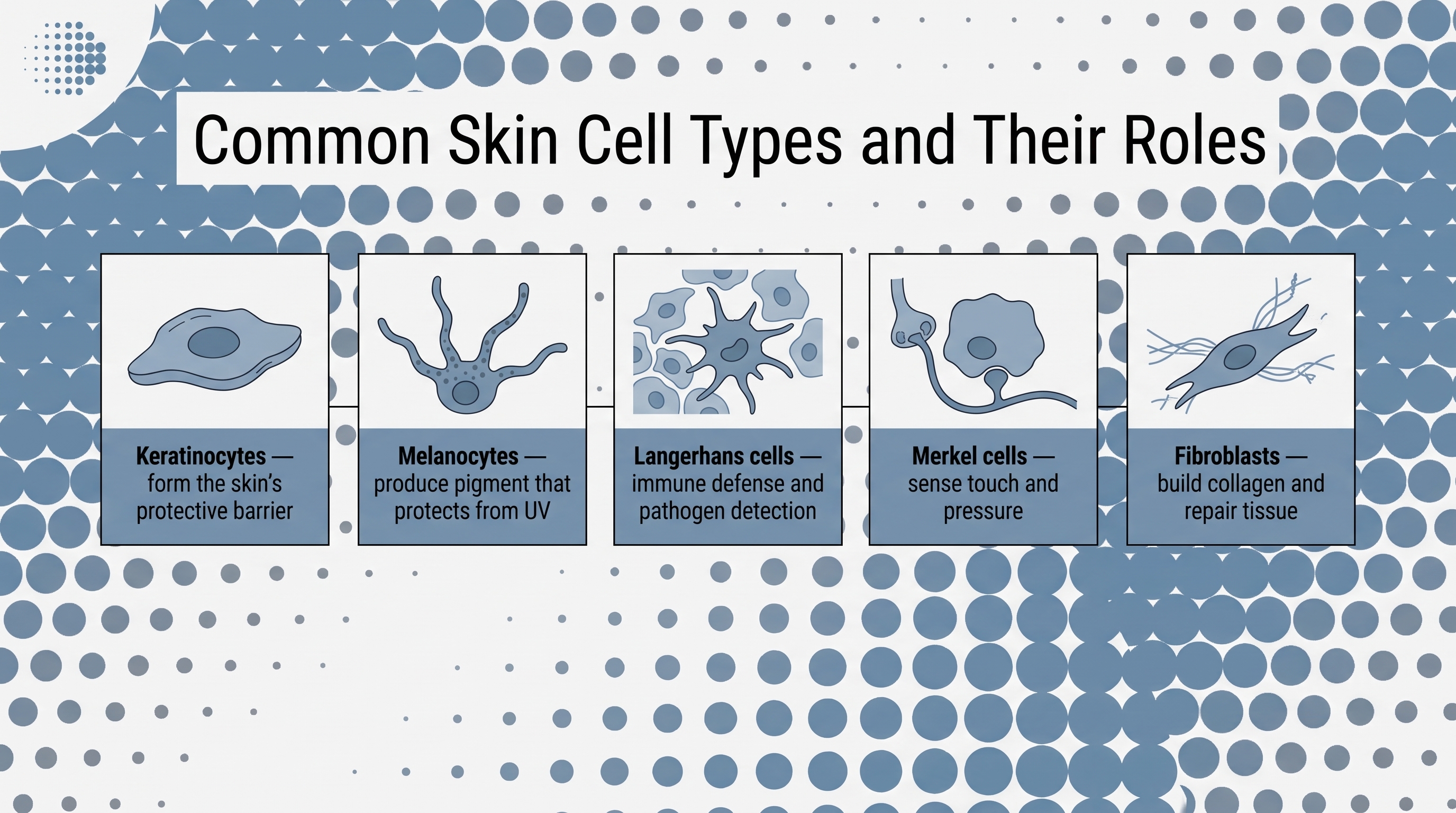
Common Skin Cell Types and Their Roles
Why the skin qualifies as the body's most complex organ Most people, if asked to name the body's most complex organ, would say …
Why the skin qualifies as the body's most complex organ
Most people, if asked to name the body's most complex organ, would say the brain. That's a reasonable instinct. But complexity isn't only about processing power. It's also about multitasking: maintaining radically different functions simultaneously within a single structure. By that measure, skin has a serious argument.
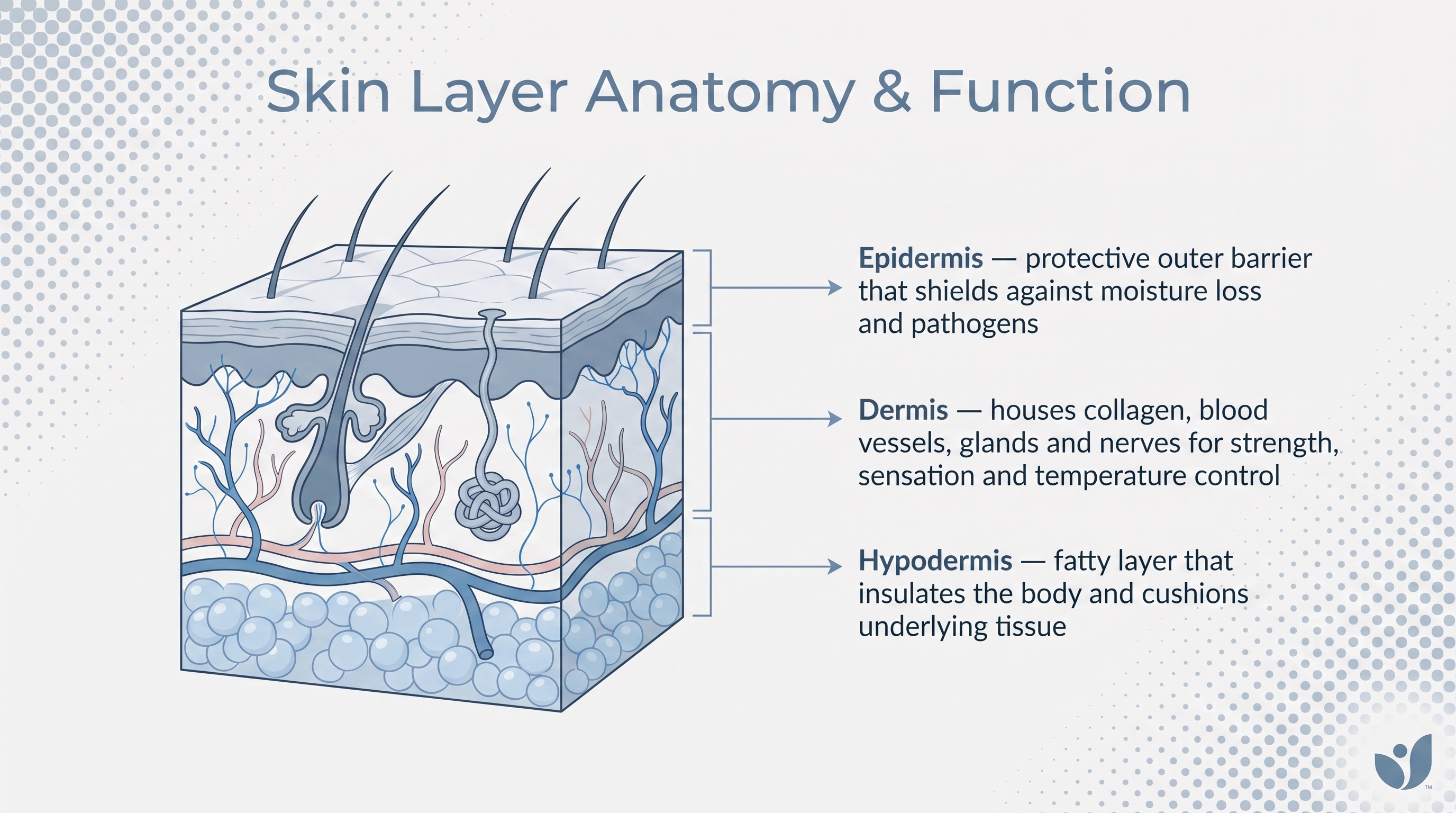
What makes skin an organ at all, rather than simply a surface, is that it contains multiple tissue types working in coordination. Epithelial, adipose, vascular, nervous. All present, all contributing, all organized within the same structure. A tissue does one thing. An organ does several, using different kinds of cells organized to do each thing well.
Skin does this across three distinct layers: the epidermis on the outside, the dermis beneath it, the hypodermis below that. Each layer has its own cellular cast, its own priorities, its own contribution. The epidermis alone hosts four specialized cell types, each performing a role the others cannot replicate. The dermis runs its own population: structural builders, immune responders, vascular components, neural elements.
None of these cells are passive. None are interchangeable. And the further you look into how they organize themselves, the stranger and more interesting it gets.
How the epidermis is organized and why cell position within it matters
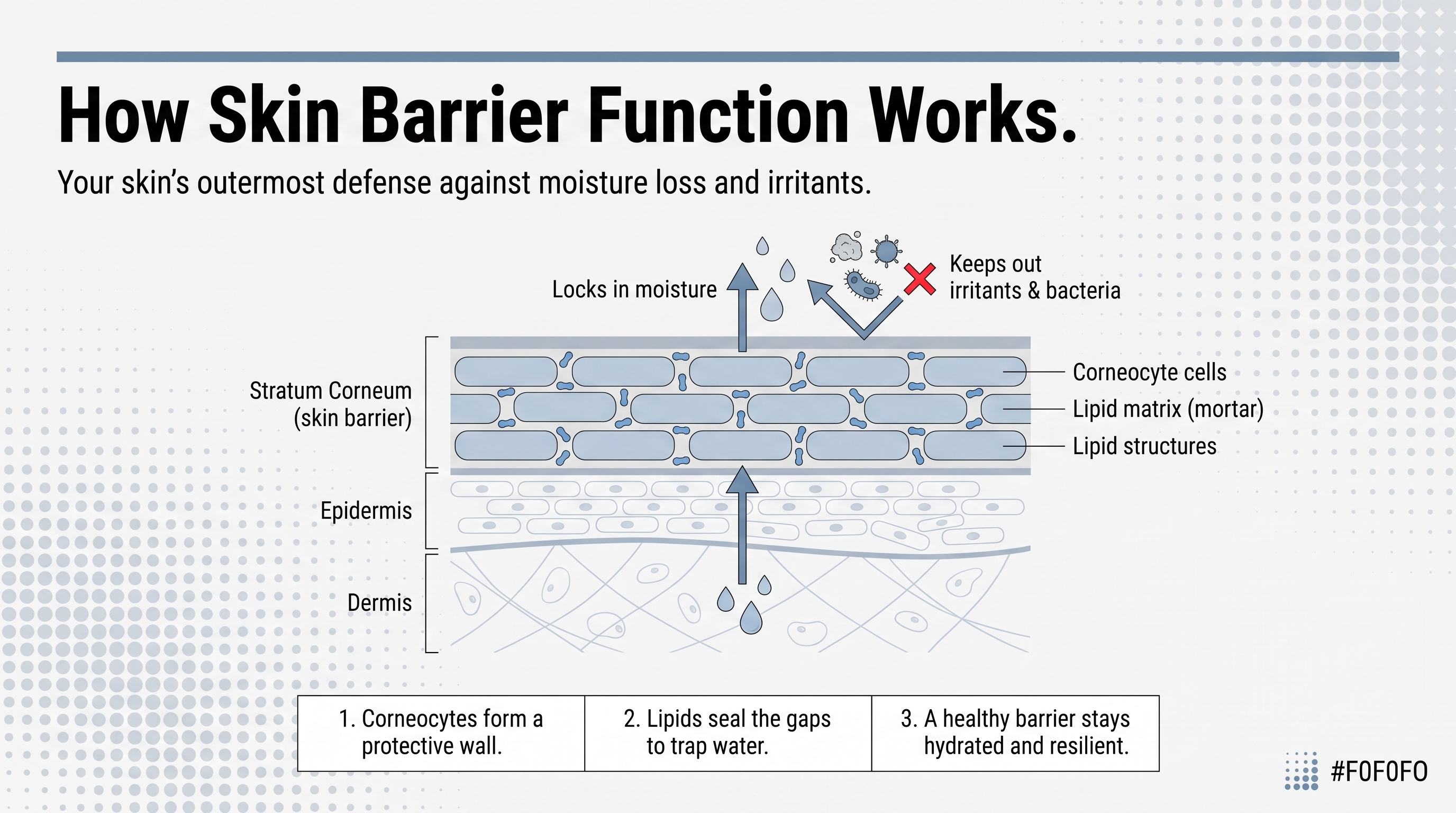
Think of the epidermis as a high-rise where the ground floor is the most productive and the top floor is, quite literally, dead. It's organized into named sublayers: the stratum basale at the deepest level, followed by the stratum spinosum, stratum granulosum, stratum lucidum (present only in thick-skinned areas like the palms and soles), and the stratum corneum at the outermost surface.
Cells are born at the base, close to the blood supply delivering oxygen and nutrients. From there they migrate upward over the course of weeks, and that journey is not just physical. As cells move away from the blood supply, they change: shape, protein expression, chemistry. By the time they reach the top, they've transformed entirely from what they were when they started.
Position within this stack is functional, not incidental. Cells doing the most active biochemical work sit near the bottom, close to nutrients. Cells forming the physical barrier occupy the top, fully differentiated and, in most cases, no longer alive. Immune surveillance and sensory reception happen toward the base and middle. The mechanical armor gets built at the top.
That logic makes the individual cell types easier to reason about as you move from the base upward, the same direction the cells themselves travel.
Keratinocytes and how they build the skin's outermost barrier
Roughly 90% of all epidermal cells are keratinocytes. That's not a plurality; that's dominance. If the epidermis is an organization, keratinocytes are the workforce, and everything else operates within the environment they build and maintain.
Their primary job is producing keratin: a tough, fibrous, water-resistant protein that accumulates as they mature and migrate upward. Keratin is what makes skin mechanically resilient. It resists heat, deflects UV radiation, slows water loss, and presents a surface hostile to bacteria, fungi, viruses, and parasites.
The differentiation journey is worth tracing concretely. Stem cells in the stratum basale generate daughter cells that begin rising through the layers. As they travel, they progressively flatten and lose access to blood supply. Molecular markers track this shift, specifically the keratin proteins expressed at each stage. Basal keratinocytes carry keratins 5 and 14. Upper-layer keratinocytes switch to keratins 1 and 10. Approaching terminal differentiation, cells begin expressing involucrin, loricrin, and filaggrin, proteins signaling the final structural transformation. By the stratum corneum, those cells are no longer alive; they're corneocytes, flattened protein-dense envelopes stacked 10 to 30 layers deep, constantly shedding from the surface. The whole cycle from stem cell to desquamation takes roughly 40 to 56 days in a healthy adult. It slows with age, which is part of why aging skin has a different texture and why it responds more slowly to both insult and treatment.
But does this mean keratinocytes are simply passive bricks in a structural wall? The picture doesn't survive scrutiny. When pathogens breach the surface, keratinocytes secrete chemokines that actively recruit monocytes, natural killer cells, T lymphocytes, and dendritic cells. They're signaling the immune system. They are participants in inflammatory response, not inert material sitting between the threat and the body.
Psoriasis illustrates this with uncomfortable clarity. Hyperproliferation of keratinocytes is one of the defining cellular features of psoriatic lesions, but the disease doesn't reduce to a keratinocyte malfunction. It's a feedback loop between autoreactive T cells and keratinocytes, each amplifying the other's response. Once you stop treating keratinocytes as passive, that loop becomes comprehensible. Before that, it doesn't.
Melanocytes and the two-cell system behind UV protection
Sitting in the stratum basale alongside keratinocyte stem cells, melanocytes represent roughly 8% of epidermal cells, approximately one for every 40 keratinocytes. Their developmental origin is entirely distinct: they migrate to the skin from neural crest cells during embryonic development, which means they are, in a meaningful sense, visitors from another lineage who took up permanent residence.
Their function is UV protection, but the mechanism is a partnership worth understanding precisely. Melanocytes synthesize melanin, package it into organelles called melanosomes, and transfer those packages to neighboring keratinocytes. The keratinocytes then position the melanin around the nucleus, arranging it like a physical shield to absorb and scatter incoming UV radiation before it reaches the DNA. One cell makes the tool; the other deploys it. Neither step alone produces protection.
The stakes here are not trivial. Epidemiological data shows that fair-skinned individuals are 20 to 70 times more likely to develop skin cancer than dark-skinned individuals. That range reflects both the protective power of melanin and the profound consequence of its absence. Pigmentation is not cosmetic in any trivial sense; it is a primary defense against one of the most prevalent carcinogens on the planet.
It is also worth considering a wrinkle introduced by research published in Scientific Reports in 2024: melanin also generates oxidative DNA damage in keratinocytes following UV exposure. I'll admit that finding took me a moment to sit with. It doesn't dismantle the framework, but it makes it more honest. The full picture of melanin's effects is more conditional than the simple shield metaphor implies, and the science is still working it out.
When melanocytes transform malignantly, the result is melanoma, one of the more aggressive skin cancers partly because of melanocytes' migratory origins and their capacity to spread. When the immune system eliminates them, the result is vitiligo. Two very different diseases, one cell type at the center of both.
Langerhans cells as the epidermis's immune surveillance layer
Langerhans cells are dendritic cells resident in the stratum spinosum, making up a small fraction of epidermal cells. Their established role is antigen capture: they sample the skin environment, migrate to regional lymph nodes, and present antigens to T cells, initiating adaptive immune responses. Allergic contact dermatitis, psoriasis initiation, the immune response to topical drugs. Langerhans cells are involved in all of it.
That role alone would justify their presence. But what if this small population is quietly doing far more than the classical model ever captured? Recent mouse-model research published through NCBI adds a dimension that hasn't fully entered mainstream clinical conversation.
That work found that Langerhans cells also orchestrate the clearance of UV-damaged keratinocytes by modulating the local chemokine environment and directing neutrophil trafficking toward compromised cells. When Langerhans cells were absent from the skin model, inflammation diminished, fewer TNF-alpha-producing neutrophils were present, and populations of DNA-damaged keratinocytes persisted longer than they should have. These cells weren't just watching for foreign antigens; they were functioning as a kind of genome-level quality control, helping identify and clear cells whose DNA had been compromised by UV exposure.
This is grounded in animal models. It's not a settled conclusion, and I'd be skeptical of anyone presenting it as one. But if it holds in human tissue, it would mean Langerhans cells contribute to cancer prevention in a more direct, immediate way than the classical antigen-presentation model ever captured.
What's already settled: a cell population comprising a small fraction of the epidermis is doing work the entire surface depends on. That proportion, tiny population carrying enormous consequence, is something you'll notice again as you move through the other cell types.
Merkel cells and why the skin needs to sense light touch
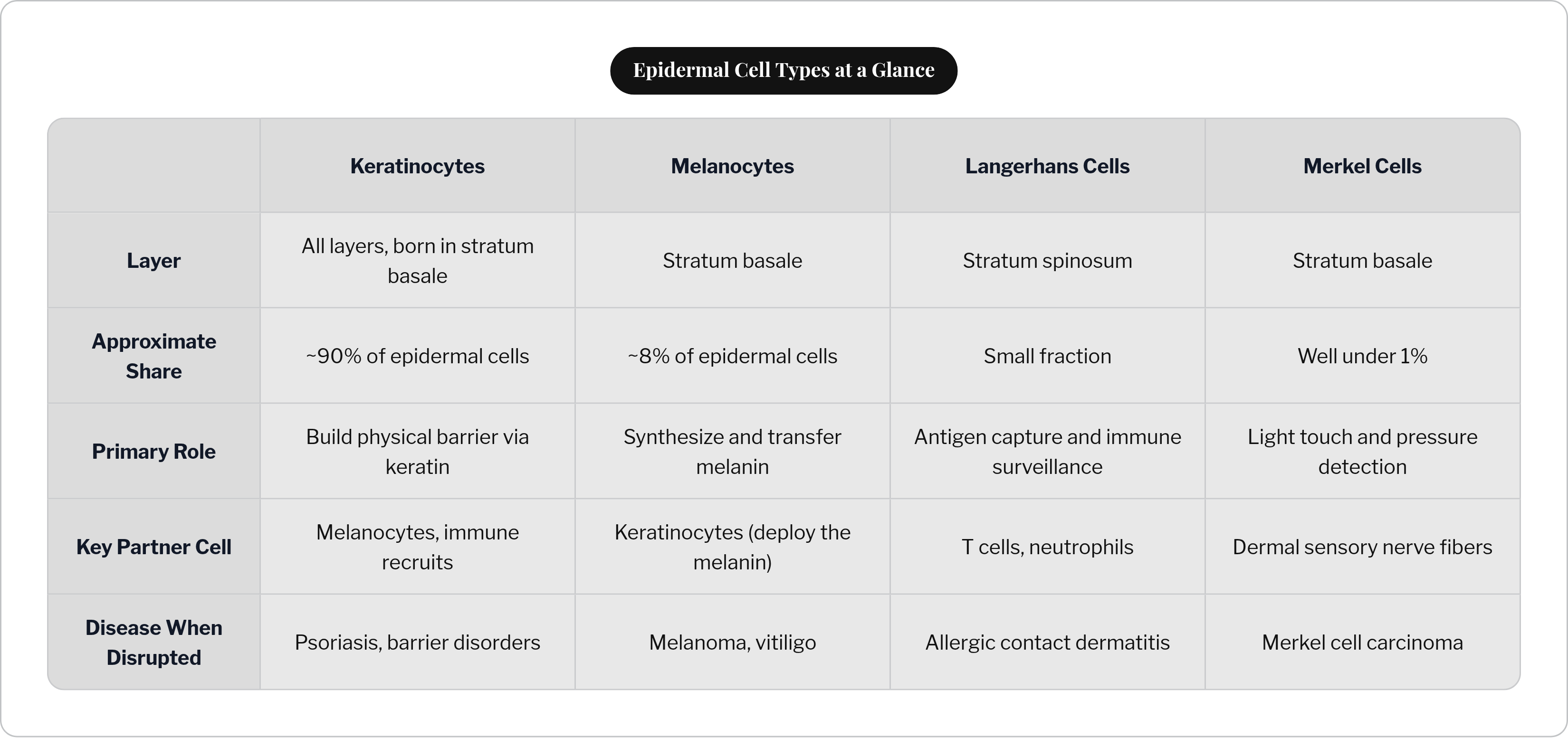
Merkel cells are the smallest population in the epidermis by a wide margin, well under 1% of epidermal cells, located in the stratum basale. They function as mechanoreceptors: specialized cells that convert light touch and sustained pressure into electrical signals traveling to sensory nerve fibers in the dermis below.
They're not floating free. Merkel cells are anchored to neighboring keratinocytes via desmosomes, the protein complexes that hold adjacent cells together. That structural integration is part of what makes them effective; they're embedded in the tissue they serve, moving with it, positioned to respond to deformation at the surface.
Their distribution across the body is not uniform. They concentrate heavily in the fingertips, where fine tactile discrimination matters most. Reading texture, feeling the edge of a surface, distinguishing smooth from rough: all of that depends substantially on Merkel cell density and function. It's one of those facts that seems obvious in retrospect but reframes how you think about why fingertip skin is structured so differently from skin on your back.
Merkel cell carcinoma is rare but aggressive, and its behavior reflects the unusual cellular identity of its origin. A cell that is simultaneously epidermal, neurological, and sensory doesn't fit neatly into the taxonomies oncologists typically work from, and the rarity of the disease can obscure how significant that is. Small populations, when disrupted, produce consequences disproportionate to their numbers. This cell type is another example of that pattern.
Moving into the dermis: what fibroblasts build and maintain
The dermis is a different environment entirely. Where the epidermis is a stratified epithelium organized around cellular migration and differentiation, the dermis is connective tissue: collagen fibers, elastic fibers, extracellular matrix, blood vessels, nerves, hair follicles, and glands, all embedded in a dense structural scaffold.
The principal architect of that scaffold is the fibroblast.
Fibroblasts synthesize collagen, elastic fibers, reticular fibers, laminin, fibronectin, and other extracellular matrix components that give skin its tensile strength, elasticity, and structural coherence. They're why skin stretches without tearing and returns to shape after deformation. They also secrete cytokines, metabolites, and growth factors that shape the local tissue environment and modulate immune responses.
Single-cell transcriptomics has revealed something earlier histological analysis couldn't resolve: fibroblasts are not a uniform population. Papillary fibroblasts, reticular fibroblasts, pro-inflammatory fibroblasts, dermal papilla cells, pericytes, mesenchymal stem cells. Each carries a distinct transcriptional signature and a distinct functional role. And the composition of that population isn't fixed. Age, anatomical location, and environmental stressors all shift which subtypes predominate, with measurable consequences for wound healing, fibrosis, and skin aging.
In scleroderma and atopic dermatitis, distinct fibroblast subtype profiles have been identified, implicating these cells as active participants in those disease processes rather than bystanders caught in someone else's inflammation. Understanding which subtypes are present, in what ratio, and what drove the shift is becoming a meaningful clinical question, not just a taxonomic curiosity for researchers.
Macrophages and mast cells as the dermis's immune and inflammatory responders
The dermis maintains its own immune presence, distinct from the Langerhans cell surveillance operating in the epidermis above.
Macrophages, referred to in dermatological contexts as histiocytes, are phagocytic cells. They engulf pathogens and cellular debris, regulate inflammation, coordinate tissue repair, and maintain homeostatic order in the dermal environment. What makes them interesting is their flexibility: they're capable of shifting between pro-inflammatory and anti-inflammatory states depending on what the tissue actually needs at a given moment. That raises an important question worth sitting with: what does it mean for a cell to read context and recalibrate its behavior? It's doing something meaningfully more sophisticated than a cell locked into one role.
Mast cells occupy perivascular areas of the dermis, sitting close to blood vessels where their effects can distribute rapidly. When activated, granules release histamine, proteases, and cytokines that drive allergic responses, inflammation, and vascular permeability changes. They're central to urticaria, to the itch of atopic dermatitis, to the early response to skin injury.
The full dermal roster is worth naming: endothelial cells lining blood and lymphatic vessels, lymphocytes patrolling the tissue, Schwann cells supporting peripheral nerve function, dermal dendritic cells, and pericytes regulating capillary function. The dermis is simultaneously a structural layer, an immune environment, a vascular network, and a neural substrate. It is not support scaffolding for the epidermis above. It is an active, immunologically sophisticated tissue conducting its own operations.
How these cell types work as a system rather than a collection of parts
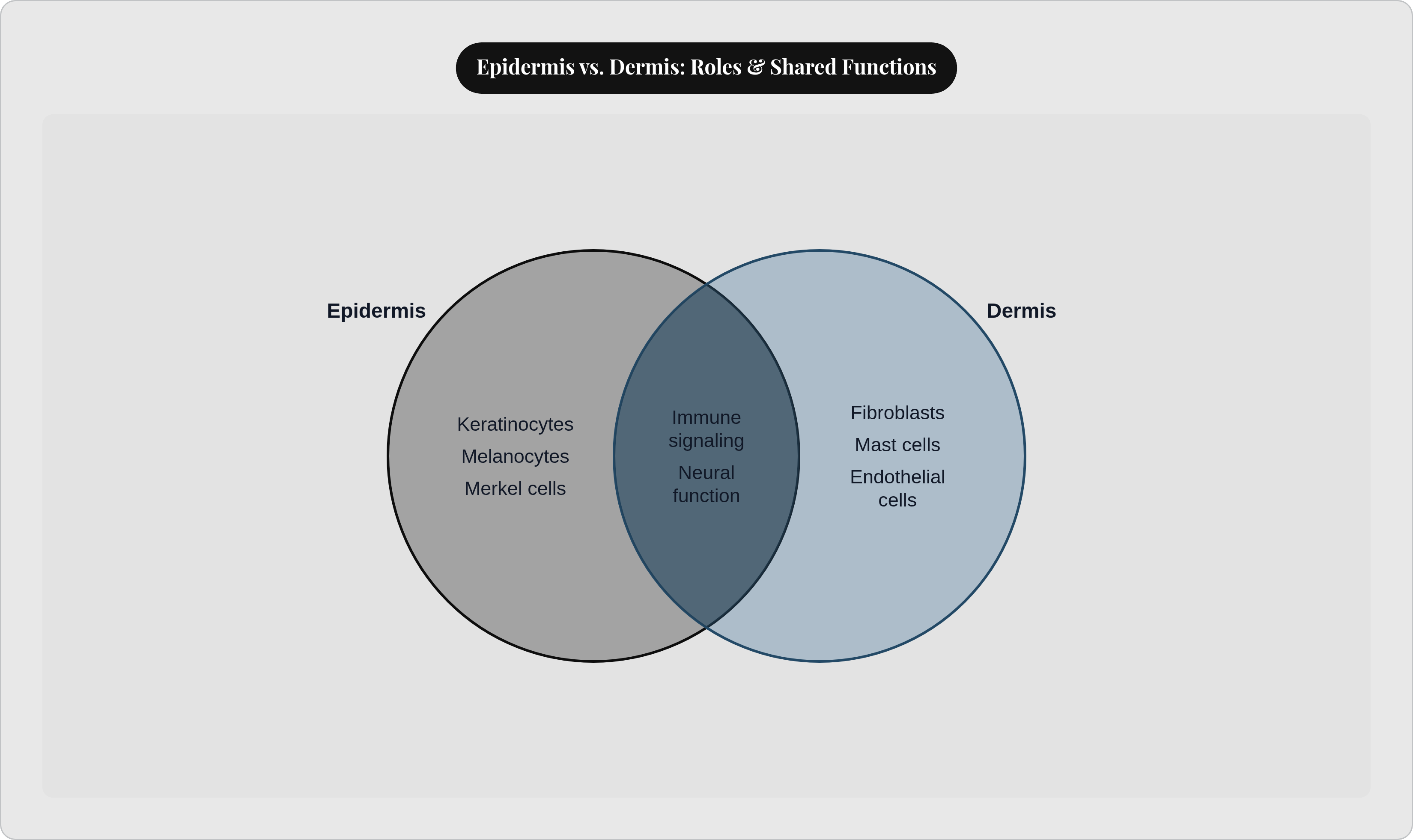
The layer-by-layer map is useful, but it can mislead if taken too literally. The epidermis and dermis are not sealed compartments. Signals, cells, and materials move across layers constantly, and function doesn't respect boundaries drawn on an anatomical diagram.
The partnerships you've traced are worth naming together, because the picture changes when you see them side by side. Melanocytes synthesize melanin; keratinocytes deploy it. Keratinocytes secrete chemokines; Langerhans cells and recruited immune cells respond. Fibroblasts build the extracellular matrix; macrophages police it, clear debris, regulate its remodeling. Merkel cells convert mechanical stimuli; dermal sensory nerve fibers receive and transmit those signals. None of these functions exist in isolation, and none of them would survive long if the adjacent cell type in this chain failed.
Disruption of one cell type doesn't stay contained; it propagates. Psoriasis is a feedback loop between keratinocytes and autoreactive T cells, but it carries dermal consequences: vascular dilation, inflammatory infiltrates, fibroblast involvement. Melanoma originates in a single cell type but carries systemic implications rooted in melanocytes' neural crest origins and migratory capacity. Fibroblast subtype shifts alter not just the structural properties of the dermis but its cytokine environment, which influences how immune cells above and around it behave.
The practical value of understanding this is direct. When a skin condition appears, identifying which cell type is implicated is the first step toward understanding why the condition behaves the way it does: why it spreads, why it persists, why a particular treatment works or fails. You can't reason about a system you haven't mapped, and the map of skin is considerably more intricate than most people ever have reason to examine.